
决赛答辩-文章数据呈现与内容补充

以下为部分文章数据的可视化呈现(部分已实现交互)









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
以下为实验思路的补充陈述
图1、本研究技术路线

图2、两个群体的亲缘关系

以下为研究进展的补充
图3、2015年以来的玉米根系GWAS和QTL研究

以下为研究结果的补充
第一部分-大田玉米根系的GWAS分析
点击此处查看DIRT平台的介绍和详情
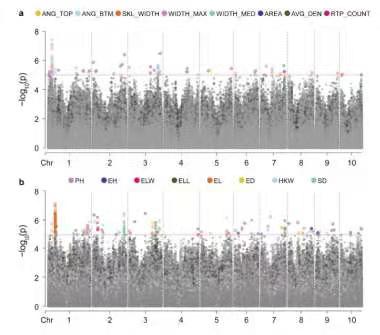
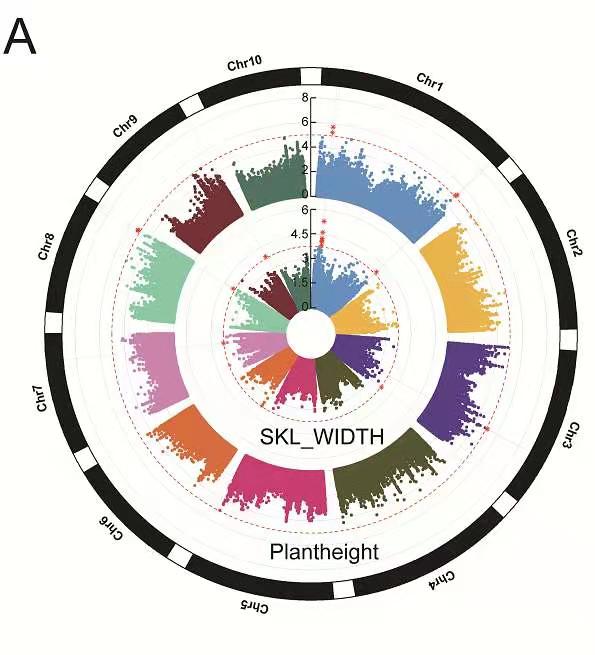
大田玉米根系的GWAS分析结果
八个根系表型性状的描述及其统计数据表

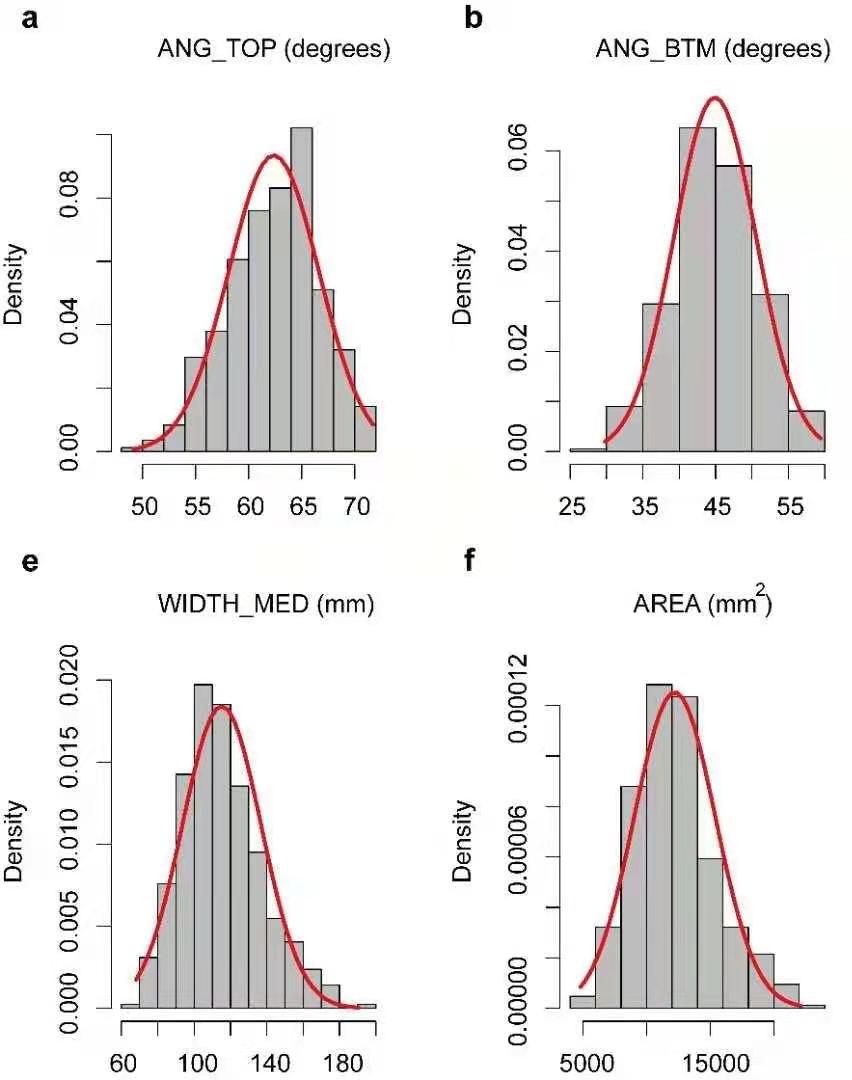
根系性状的概率分布直方图

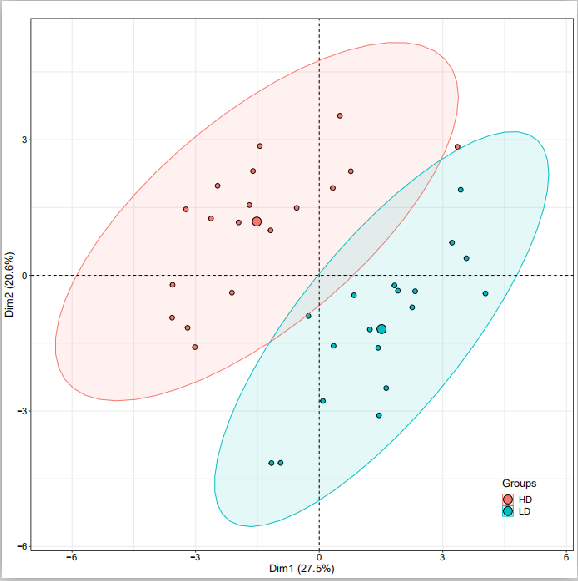
根系构型性状和地上部性状的主成分分析

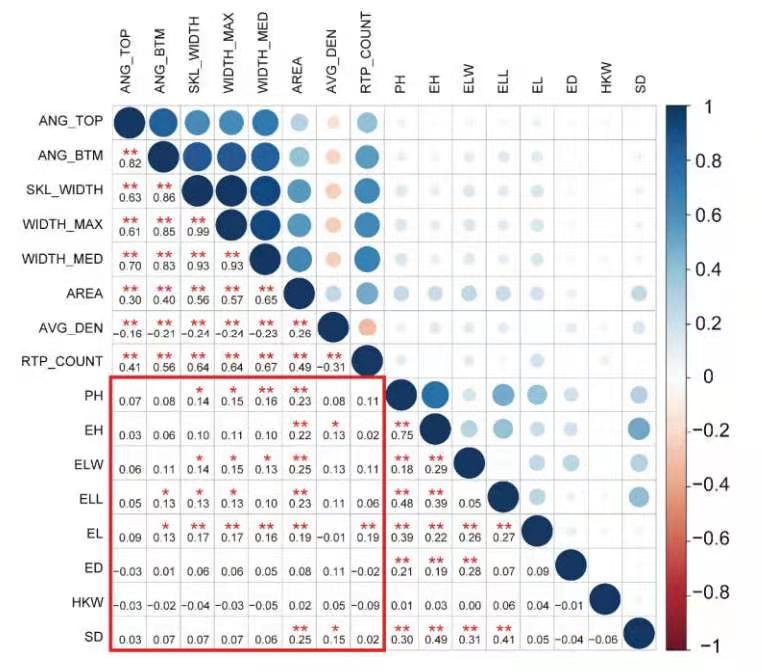
根系构型性状和地上部性状的相关性分析

不同亚群根系性状的差异分析
-scaled.jpg?fit=1000%2C529 "不同亚群根系性状的差异分析")
地上部性状与根系性状的遗传相关

根系和地上部性状关联 SNP 和候选基因数目

根系性状候选基因的共表达网络分析

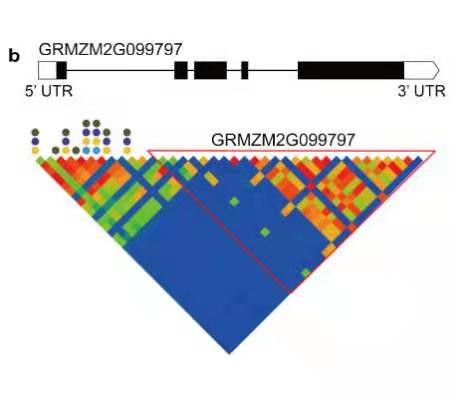
候选基因 GRMZM2G099797 连锁分析及单倍型分析

玉米根系性状关联的高优先候选基因

至此,我们挖掘了4个高优先的根系构型候选基因
第二部分-不同耐密性玉米品种地上部和根系性状的协同响应
地上部和根系性状的方差成分分析

不同种植密度下地上部和根系性状的差异分析

不同品种玉米的产量均值和分组

不同玉米品种亚类的性状差异比较

不同玉米品种类型的干物质分类差异

不同种植密度下的各个性状与产量的相关系数

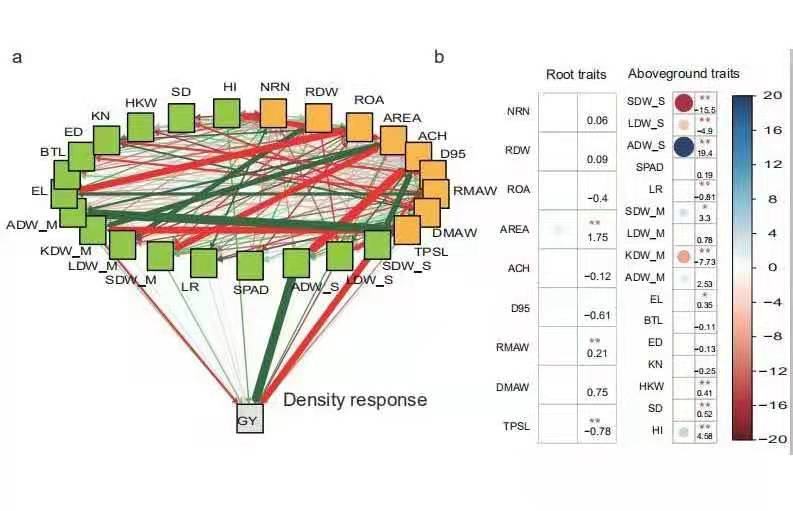
地上部性状与根系对籽粒产量的贡献

高密下理想的地上部根系协同构型

补充内容-附件
附图
1、Histogram of distribution of eight root traits.
2、Principal component analysis of root and aboveground traits.
3、Manhattan plot of eight root traits analyzed by GWAS
4、Manhattan map of aboveground traits analyzed by GWAS.
5、Co-expression network analysis of root candidate genes.
6、GWAS identification of high-priority genes for variation in maize root traits.
7、Figure legends of Supplementary Figures
点击此处下载
附表
Table S1. The number of significant SNPs and candidate genes associated with eight root traits.
Table S2. SNPs significantly associated with eight root traits and the candidate genes within 50 kb up- and downstream of each unique significant association SNPs.
Table S3. The number of significant SNPs and candidate genes associated with eight aboveground traits.
Table S4. SNPs significantly associated with eight aboveground traits and the candidate genes within 50 kb up- and downstream of each unique significant association SNPs.
Table S5. List of high-priority candidate genes generated from co-expression network analysis.
Table S6. Information from the maize lines in the panel, including the pedigree and subpopulation.
Table S7. The phenotypic data of eight root traits.
Table S8. Description of eight aboveground traits.
Table S9. The phenotypic data of aboveground traits used in this study.
点击此处下载
18个杂交种信息
附件文献
联系方式
Email :wubin_rush@163.com
Wechat:w1279868039